
Floral Identity
In 1991, Enrico S. Coen & Elliot M. Meyerowitz proposed the ABC Model as a means of explaining how floral whirls develop in Arabidopsis thaliana and snapdragons (Antirrhinum majus).
With the exception of monocotyledonous species and basal angiosperms, the flowers of most Eudicot species are composed of four different floral organ types:
- Sepals
- Petals
- Stamens (anther and filament, otherwise known as the androecium [male])
- Carpels (stigma, style and ovary, otherwise known as the gynoecium [female])
These four components are all arranged in individual whorls around the meristem with the carpels being innermost. In some species there is no distinction between sepals and petals, and these are called tepals - as seen in Amaryllis for instance.
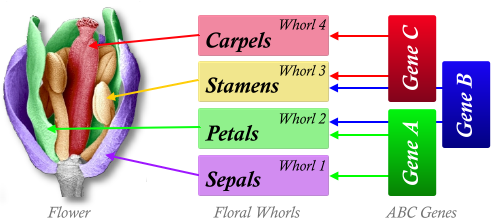
In both Arabidopsis and Antirrhinum, the genes which control each whorl are known. The genes are classified by their function and are referred to as homeotic genes:
- 'A' genes control the sepals
- 'A' and 'B' genes in combination control the petals
- 'B' and 'C' genes in combination control the stamens
- 'C' genes control the carpels
In the floral meristem, the 'A' genes are read first and produce sepals (whorl 1) from the floral meristem. Secondly the 'A' and 'B' genes are read together and petals are produced (whorl 2), then the stamens (whorl 3; 'B' and 'C' genes) and then the carpels (whorl 4; 'C'), by which time there are no active meristematic tissues left.
The four whorls are created in this way because of the 'A', 'B' and 'C' gene groups working in this order. If a gene is removed or deactivated, then changes can happen.
An important aspect of this system is that it appears that the 'A' and 'C' type genes block the action of each other so that the two cannot work together. This is thought to stop the continuation of the flower. In some mutants of Antirrhinum (such as the floricaula homozygous mutant), an inflorescence forms in the fourth whorl and no actual floral organs form. (This is because the inflorescence meristems never develop into floral meristems - for more info see Floral Reversion).
Across a number of species it is possible to identify the floral organ homeotic genes, and classify them as having A, B, or C function. In Arabidopsis, the homeotic 'A' genes are APETALA1*,APETALA2, and LEUNIG. The 'B' genes are APETALA3* and PISTILLATA* and the 'C' gene (there is only one) is AGAMOUS* (Ma & dePamphilis 2000). (Those marked * are MADS Box genes).
Studies in a wide range of species have subsequently provided support for the ABC model, with anomalies being attributed to idiosyncrasies of individual species. In partcular, similar classes of mutant have been described in both Arabidopsis and Antirrhinum, suggesting that the regulation of organ identity is highly conserved. Mutants with defects in the second and third whorls, which result in the homeotic conversion of petals to sepals and stamens to carpels have defined the B-function and include the Arabidopsis pistillata (pi) and apetala3 (ap3) mutants and the Antirrhinum globosa (glo) and deficiens (def) alleles. However, some experiments have yielded results that are not entirely compatable with the ABC model and suggest a further degree of complexity.

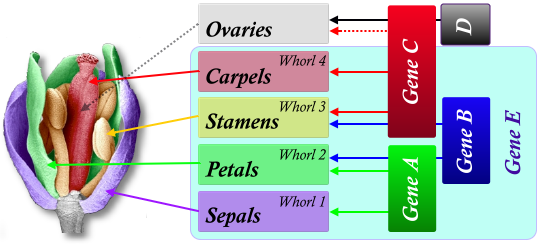
One of the challenges with the ABC model is that one would expect that one of the A, B, or C genes, if constituatively expesssed in vegatative issue, would cause that tissue to become floral. Unfortunately such experiments with transgenic plants has shown this not to be the case, indicating that ABC genes only function within a pre-defined floral context. As a result, the ABCE Model has been developed.
The isolation of novel floral mutants in Arabidopsis (and other species) has led to an expansion of the ABC model to include the D- and E-functions. (The D-function specifies ovule identity in combination with members of the C-function group of genes).
Studies on tomato and petunia have revealed the TM5 and FBP2 MADS box genes, respectively. When silenced, the resulting plants have an appearance that suggests a reduction of B- and C-gene function. When SEPALLATA 1 (SEP1),SEP2 and SEP3 genes were found in Arabidopsis and silenced, they also showed a similar appearance whereby all floral organs were replaced with sepals. Expression analysis of these genes, and related genes from Antirrhinum (DEFH72 and DEFH200), revealed that they are expressed after the floral meristem identity genes, but before the onset of the organ identity genes in all whorls of the flower. In essence, the role of these E-genes is to establish the floral context in which the organ identity genes can function.
Additionally, Causier et al. (2010) question the very role of A-genes ascribed to them in the classical ABC model. In particular, they note the role of A-genes extends outside the flower (unlike B- and C-genes) but note that the A-genes are required for the later activation and regulation of the B- and C-functions and therefore results in the establishment of B- and C-function boundaries.
Hence, the new ABCE model for floral development could be summarised as such:
- 'A' genes control the sepals and activation of 'B' and 'C' genes
- 'A' and 'B' genes in combination control the petals
- 'B' and 'C' genes in combination control the stamens
- 'C' genes control the carpels
- 'C' and 'D' genes control ovule identity and development
- 'E' genes establish a floral context in which the 'A', 'B' and 'C' genes can function
Many discoveries have been made in the last 30 years about floral identity and how flowers are "built".
Double flowers
Double flowers are commonplace and have been actively selected by humans for millenia; think of double roses, carnations and petunias. The genetic basis of the double flower trait was recently deciphered in ornamental plants belonging to the Rosaceae family, where the PETALOSA (PET) gene was found to play a crucial role in causing extra whorls of petals to form. Gattolin et al. (2020) used a genetic modification system called CrispR-Cas9 to induce changes in the miR172 target site of tobacco PET genes. The resultant plants produced extra whorls of petals.
Next: Floral Reversion